Research
We combine nonlinear dynamics, spatial ecology, and applied mathematics to understand and predict the dynamics of spatially extended systems. Across diverse ecosystems and scales, we seek general principles governing how spatial organization emerges, evolves, and responds to environmental change.
How do spatial patterns originate, interact with their environment, and change?
Pattern formation • Nonlinear dynamics • Self-organization


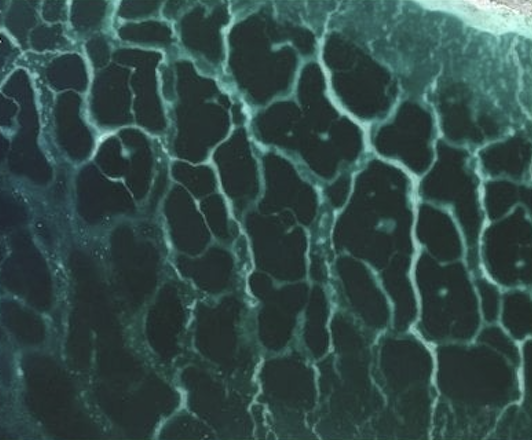
From left to right: (A) regular vegetation patterns in drylands; (B) microbial communities forming cone structures in Antarctic lakes; (C) reticulated coral reef patterns.
We develop mathematical models to understand how interactions among organisms, resources, and the physical environment generate self-organized spatial patterns. By comparing diverse ecosystems—including dryland vegetation, biological soil crusts, Antarctic microbial communities, coral reefs, coastal wetlands, and soil redox patterns — we seek general mathematical principles governing pattern formation in living systems and their ecological and evolutionary consequences.
MODEL SYSTEMS
-
Dryland vegetation and biological soil crusts
Funded by NSF-DEB | Collaborators: Yufang Jin, Rachata Muneepeerakul, Caroline A. Havrilla, Yu Zhang -
Antarctic microbial communities (modern stromatolites)
Funded by NSF-OPP | Collaborator: Dawn Sumner
TOOLS
Reaction–diffusion models Partial differential equations Stability analysis Computational modeling
How do evolution and spatial organization shape one another?
Eco-evolutionary dynamics • Biogeomorphology • Landscape evolution

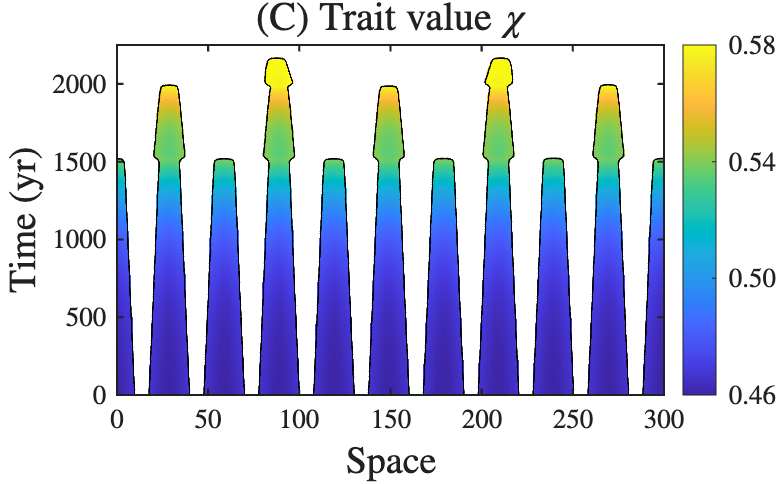
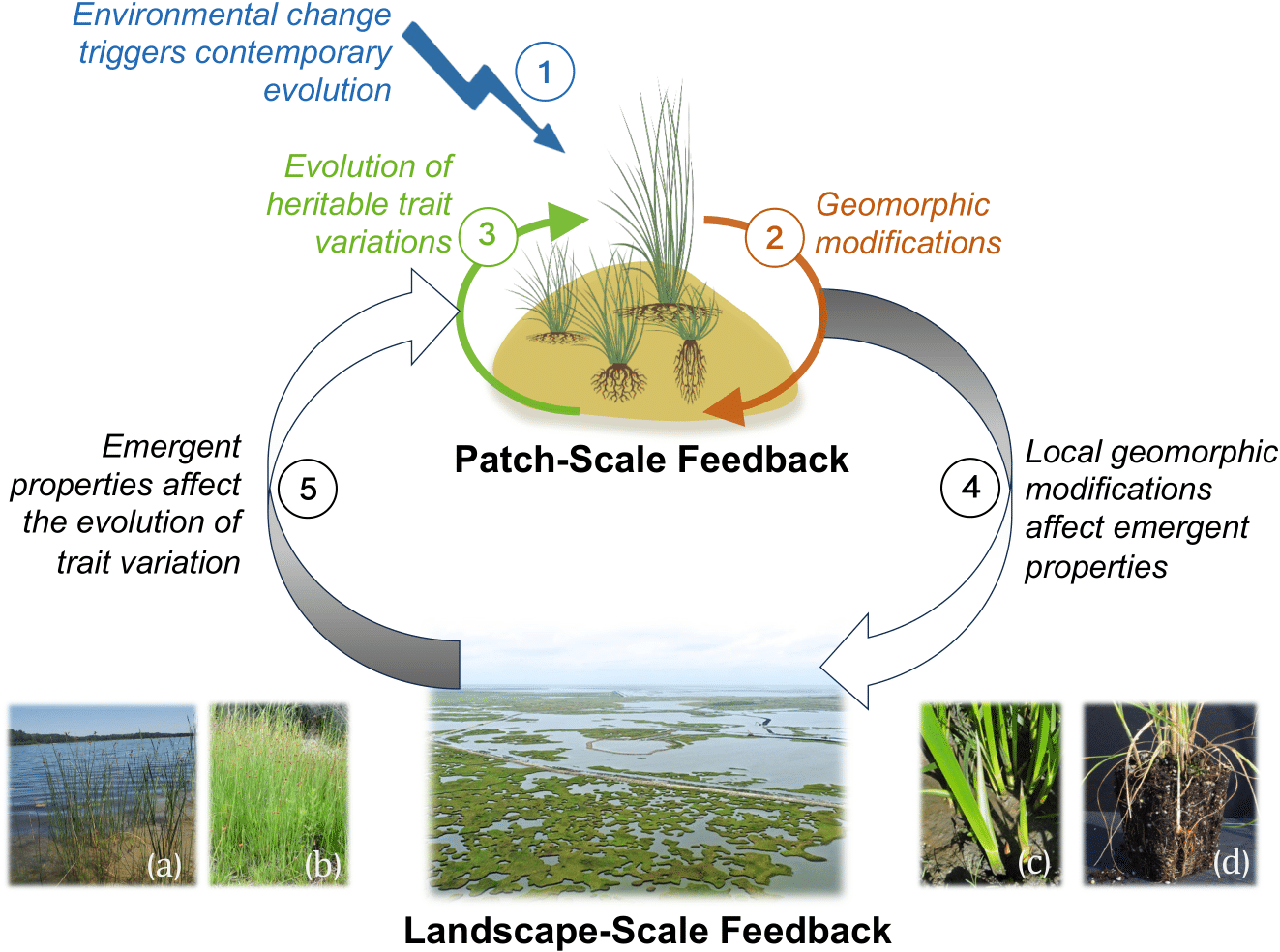
From left to right: (A) plant traits shaping the morphology of riverine landscapes; (B) spatial self-organization inhibiting adaptive evolution (work under review); (C) eco-evolutionary feedbacks between organisms and landscapes (read more in TREE (2024)).
Organisms continuously modify the landscapes they inhabit, while changing landscapes reshape ecological interactions and natural selection. We develop mathematical theory to understand how ecological, evolutionary, and geomorphic processes interact to drive the emergence and evolution of spatial organization. By integrating eco-evolutionary dynamics of organisms with evolution of landscapes, we seek general predictive principles governing the long-term dynamics of both landscapes and the organisms that inhabit them.
MODEL SYSTEMS
-
Riverine and biogeomorphic landscapes
Recent work: TREE (2024), The American Naturalist (2022) - Dryland vegetation
TOOLS
Eco-evolutionary theory Landscape evolution models Partial differential equations Computational modeling
How can mathematics and AI improve environmental prediction?
Scientific AI • Physics-informed machine learning • Earth system prediction

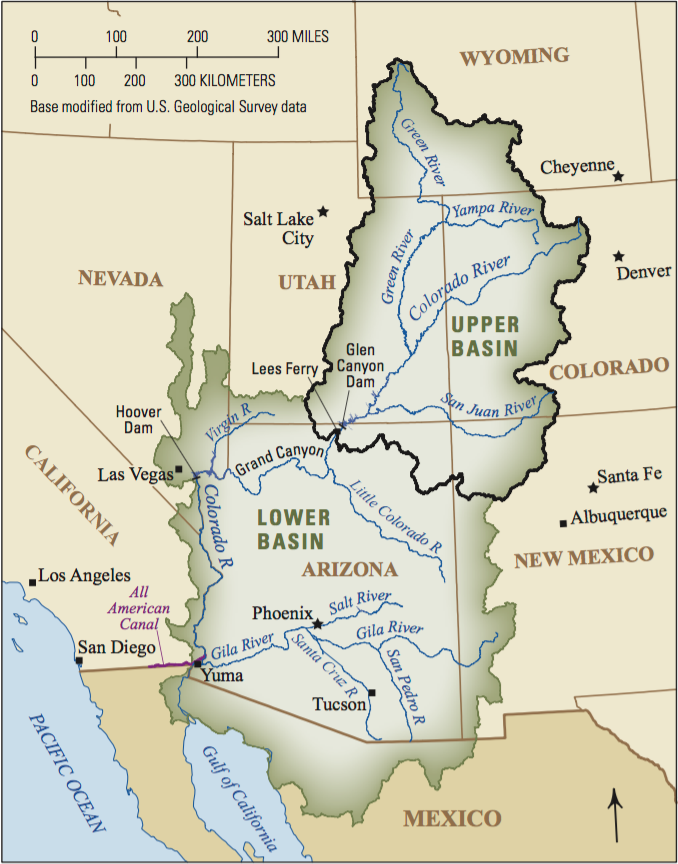
From left to right: (A) colorado river; (B) map of colorado river basin; (C) architecture of temporal transformer.
Mechanistic models provide scientific understanding but are often limited by incomplete process knowledge, whereas machine learning can uncover patterns from large datasets but often lacks physical interpretability. We develop physics-informed artificial intelligence that integrates mathematical models with modern deep learning to improve environmental prediction while maintaining scientific interpretability. Our current work focuses on hydrological forecasting through collaborations spanning applied mathematics, earth science, computer science, and ecology.
MODEL SYSTEMS
-
Colorado River Basin
Funded by the DOE GENESIS | Partners: EnviTrace LLC (Mikhael Semaan, Velimir Vesselinov), UC Davis (James Crutchfield), Oak Ridge National Laboratory (Forrest Hoffman), and Lawrence Berkeley National Laboratory (Adam Rupe)
TOOLS
Physics-informed AI Scientific machine learning Information theory Community Earth System Model (CESM)
How will biodiversity reorganize under global environmental change?
Climate change • Biodiversity • Species distributions
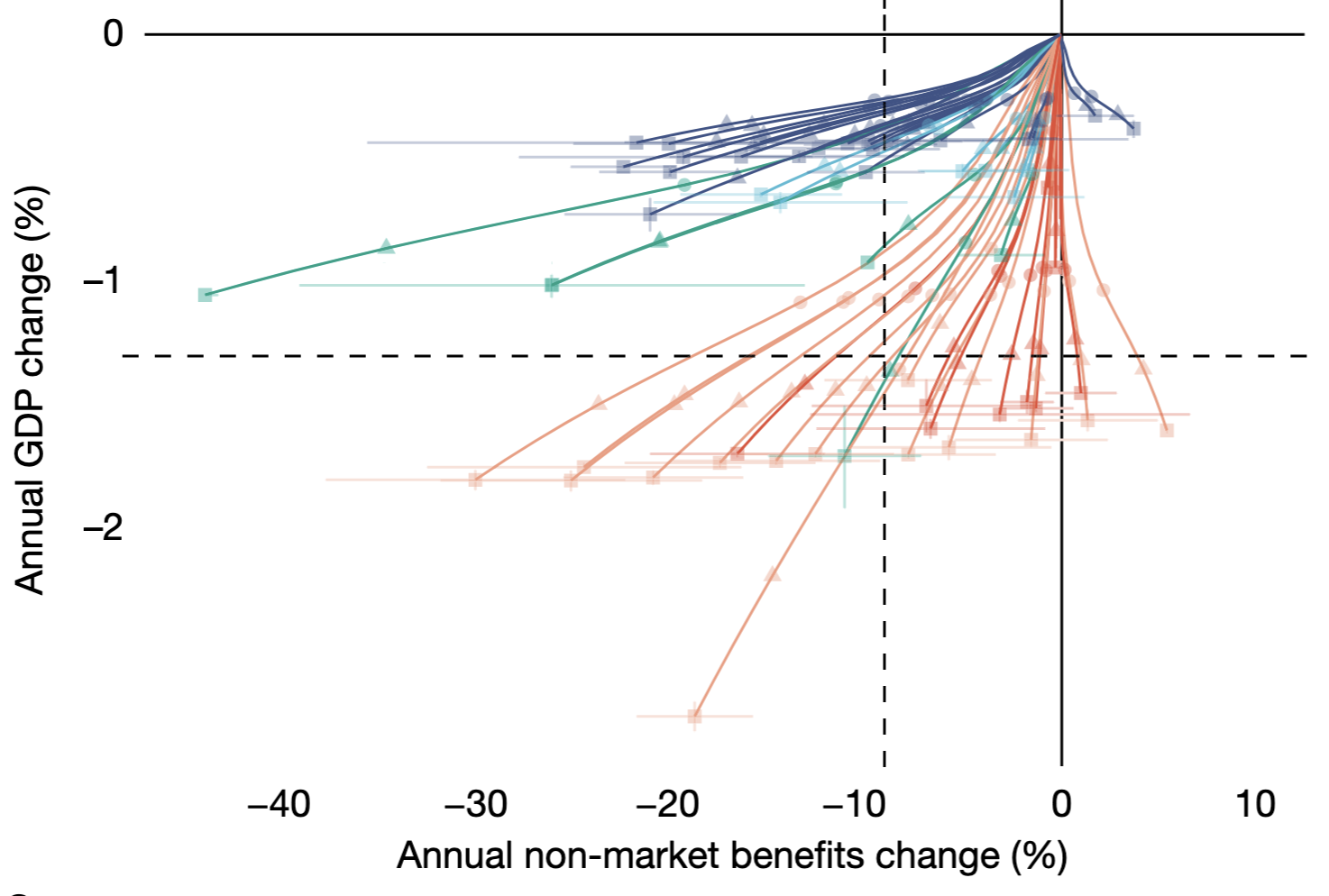
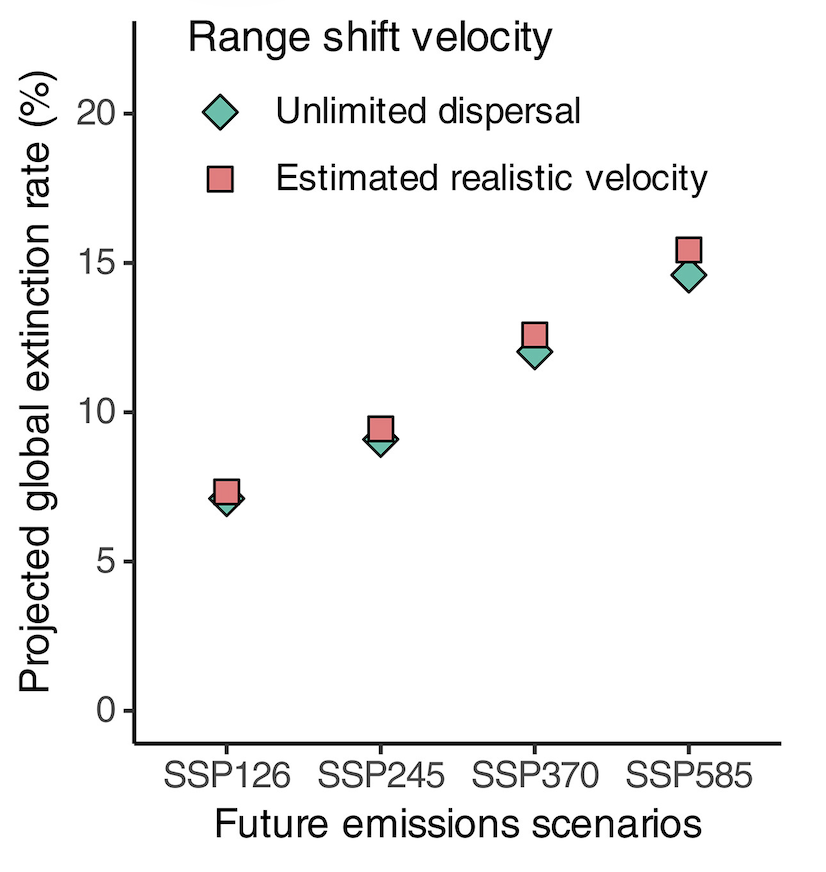

From left to right: (A) trajectories of the annual change in market and non-market benefits for the 57 countries and regions by 2100 (read more in Nature (2023)); (B) dispersal limitation plays a surprisingly small role in projected global plant extinction by 2100 (read more in Science (2026)); (C) reporting of our work by the Associated Press.
Climate change is reshaping the geographic distributions of species worldwide, creating novel communities and transforming ecosystem functioning. We develop large-scale species distribution models for ecological forecasting to understand how dispersal, environmental change, and landscape heterogeneity jointly determine future biodiversity patterns. By integrating ecological forecasting with economic and policy frameworks, we also evaluate the societal consequences of biodiversity change.
MODEL SYSTEMS
-
Global terrestrial plants and biomes
Funded by NSF-DEB | Collaborators: Francis Moore, Marc Conte
Recent work: Science (2026), Nature (2023), JUE (2023)
TOOLS
Species distribution models (SDMs) Integrated assessment models (IAMs) Macroecology